Filo Mollusca:
Os moluscos representam um dos maiores e mais diversos filos de todo o reino animal. Junto aos artrópodes (insetos), estes possuem o maior número de animais descritos de todo o reino animal (Hickman, 1961). Tanto os moluscos extintos como os vivos compreendem cerca de 11% de todas as espécies animais que já existiram em todo o curso da história da terra.
O nome Mollusca é derivado da palavra em latin Molluscus, que significa mole. Uma das características mais distintas é um corpo interno mole que contrasta com uma concha exterior dura.
Moluscos são um diverso grupo de organismos que divergem de um ancestral comum chamado ‘Arquimolusco’. Sua morfologia era composta de uma concha do tipo prato (apenas uma capa), um pé macio por onde se arrastava, um simples canal alimentar e um manto. Diferente dos moluscos modernos este organismo era segmentado.
Por todo o filo a estrutura do corpo é similar. Todos os organismos das diversas classes possuem uma cabeça para alimentação, um pé ventral e muscular usado para movimentação e víceras que compõem o resto do corpo acima do pé. As víceras também contêm orgãos internos que acomodam a cavidade do manto de onde todo o lixo é excretado.
Os moluscos são divididos em 8 (oito) classes, mas apenas 3 (três) possuem importância geológica. Estes são os bivalves, os cefalópodes e os gastrópodes.
Classes
Classe Monoplacófora
Em latin monoplacophora significa ‘que possue apenas uma placa’.
Acreditavasse que estivessem extintos a pelo menos 400 millhões de anos, quando foram encontrados animais vivos em 1952 de Neopilina galatheae Lemche, 1957 – na costa do méxico a mais de 3500 metros de profundidade. Foi como encontrar um elo perdido entre os moluscos e os anelídios. Existem apenas 6 (seis) espécie conhecidas ainda viventes. São uma importante fonte de estudos evolucionários já que possuem um corpo que é base para a evolução de todos as outras classes de moluscos.
Embora tenham sido muito comuns no período Paleozóico em águas rasas, hoje não podem mais ser encontrados nestas áreas. O descobrimento de espécies de animais considerados extintos, nos leva a rever estas considerações. As águas rasas são fisicamente mais variáveis do que as águas profundas onde salinidades, pressão e temperatura são mais estáveis.
Ecologia
Considerados nada comuns em coleções particulares, devido as profundidades onde vivem, algumas espécies vivem em profundidades relativamente rasas. Na costa da Itália, espécimens de Veleropilina podem ser encontradas em profundidades de 180 metros. Mas a grande maioria ainda vive em profundidades abissais.
Estas profundidades não permitem que animais vivos sejam observados, então pouco se sobre a sua ecologia.
Observando-se a concha, baixa, que produz pouca resistência e o formato do pé, pode-se concluir que são animais adaptados para viver em águas rasas e de forte movimento. Talves este seja um legado dos tempos em que estes animais viviam em águas costeiras. Várias espécies têm sido encontradas em áreas com nódulos de ferro e mangânes, cascalho de coral, basalto e fundos lodosos ou rochosos.
Estrutura
São animais pequenos, que variam de 3 até 35 mm. Sua concha em forma de colher é composta de 3 camadas: perióstraco, uma camada prismática e uma nacarada.
Alimentação
Acredita-se que os Monoplacóforos alimentem-se de espólios. Há informações de vários tipos de alimentos como: protozoários, diatomácias, foraminíferos e esponjas.
Classe Polyplacophora
A Classe Polyplacophora, também popularmente chamada “Chiton” representa moluscos achatados dorso-ventralmente com oito valvas articuladas dorsalmente formando uma concha; um cinturão (podendo ser referido eventualmente como perinoto) pode variar de estreito a largo, podendo estar esculturado por diferentes estruturas (região dorsal, borda e ventral) desde espículas com ou sem articulação a escamas lisas ou fissuradas e filamentos branquiais localizados na cavidade do manto, podendo ocupar de ¼ a ¾ da cavidade do manto que está recoberta pelo pé.
A classificação desses organismos ainda é muito confusa, sendo que é baseada na morfologia (valvas (exposição e escultura), cinturão (de acordo com sua largura), filamentos branquiais e mais recentemente pelas estruturas encontradas no ovo). Suas valvas podem ser classificadas de três tipos distintos: valva I (cefálica), valvas II a VII (medianas ou intermediarias) e valva VIII (anal); as brânquias podem classificar o organismo como holobrânquial (filamentos brânquias ocupando ate ¾ da cavidade do manto ou merobrânquial (filamentos branquiais ocupando ¼ da cavidade do manto).
Análise morfológica da Classe Polyplacophora
O cinturão pode ser ornamentado dorsalmente por espículas hialinas (articuladas como o gênero Chaetopleura ou não articuladas como o genero Hanleya), estruturas semelhantes a bastões (lisos – Leptochiton ou sulcados – Acanthopleura), escamas (lisas – Chiton e fissurados – Ischnochiton).
As valvas nestes representantes dos moluscos são sem duvida, as informações morfológicas mais representativas para a classificação do grupo, a primeira característica observada é a exposição da valva no cinturão, onde, pode estar na sua maioria exposta como em Leptochiton pode estar parcialmente exposta como em katharina ou totalmente recoberta como em Cryptochiton.
A valva cefálica (Valva I) pode apresentar forma semi-elíptica como no gênero Mopalia e elíptica como no gênero Leptochiton, dentes de inserção (ou pranchas de inserção) podem ser contínuos como o gênero Hanleya ou separados em dentículos menores como Chaetopleura. Ainda abordando os dentes de inserção eles podem ser sulcados longitudinalmente como Ischnoplax ou lisos como Chaetopleura.
As valvas intermediarias são as estruturas com mais modificações encontradas. A relação da escultura geralmente é muito difícil se concluir um padrão, devido ao fato que a escultura presente, não segue o mesmo padrão por toda a valva, por exemplo: a escultura na área central em grande parte dos casos não é a mesma da área lateral (como é o caso de Chaetopleura angulata), que por sua vez, em algumas espécies, segue o padrão da valva por inteiro (como é o caso de Leptochiton darioi). O ápice pode ser bem definido como em Ischnochiton striolatus, pouco definido como em Calloplax janeirensis ou inconspícuo como Leptochiton darioi. Usando como forma de comparação à linha lateral (linha que divide a área central = área pleural + área jugal) pode observar que pode esta visível como em Ischnoplax incurvata ou pouco visível como em Leptochiton asellus. A área lateral pode estar elevada como é o caso de Ischnoplax pectinata ou não como é o caso do gênero Halneya. As áreas jugal e pleural só se modificam de acordo com a escultura. Apófise (ou lamina sutural) é uma estrutura característica das valvas intermediarias e anal, sendo esta, importante na sua forma como sendo mais alargada como é o caso do gênero Acanthopleura ou então mais comprida, como é o caso do gênero Acanthochitona.
A valva anal apresenta uma dilatação denominada mucro, de acordo com a posição dessa estrutura, podemos chegar ao nível de grandes grupos, como é o caso do gênero chiton que na sua maioria apresenta mucro elevado, o gênero Ischnochiton apresenta mucro mediano e o gênero Pilsbryellla.
Dados Ecológicos sobre a Classe Polyplacophora
Hábito alimentar micrófago (raspadora de rochas) na sua maioria, os chitons se alimentam do que estiver incrustado em substrato consolidado, mas alguns poucos exemplares da classe, apresentam hábito alimentar predador.
Encontrado na sua maioria em rochas (face oculta de seixos no substrato), como também em fendas de rochas, em algas, esponjas e conchas, mas, algumas poucas espécies, podem ser observadas em fundos de areia lodosa. Podem ser encontrados dos 0 aos 1000m de profundidades.
A reprodução na sua maioria é externa (liberação dos gametas no meio), podendo apresentar cuidado parental (manutenção dos ovos na cavidade do manto). Após a fecundação, a larva apresenta um estágio planctônico (superfície da água) e depois, com a modificação da larva em um novo indivíduo, a larva cai pra região bentônica (fundo do oceano).
Poucas espécies entre os chitons podem apresentar simbiose, entre estes, Cryptochiton stelleri o maior representante na classe até o momento, pode apresentar entre os filamentos branquiais uma espécie de carangueijo (carangueijo ervilha Opisthomus transverse) e um poliqueta (Acholoe vittata).
Curiosidades sobre a classe Polyplacophora
Entre as principais curiosidades do grupo podemos citar sua importância como base alimentar para alguns nativos de determinadas regiões, como é o caso de Acanthopleura granulata que já foi observada sendo comida pelos nativos de Bermudas e Antilhas e Cryptochiton stelleri e Katharina tunicata que servem como base de alimento aos nativos do Alasca.
Classe Aplacófora
Aplaco-phora, que significa “que não possui concha”, são raramente coletados sem grande esforço e gastos. Por não possuírem uma concha, não são integrantes de coleções particulares.
Estes animais são mais parecidos com vermes, com uma cobertura brilhante composta de inúmeros espinhos ou escamas formados de carbonato de cálcio, também conhecidos como escleritos. Cada espinho é espelido de maneira extra celular como nos poliplacófora. O processo se inicia com o encapsulamento de uma célula individual que cresce através da cutícula. Estes espinhos são compostos de aragonita, com excessão das espécies de Notomenia, que não possuem Carbonato de Cálcio.
Abaixo desta capa está o que literalmente o identifica como um molusco, uma rádula (embora algumas espécies não possuam). A rádula não parece ter a mesma função de raspador, com excessão das espécies da família Prochaetodermatidae. Existe uma boca anterior e posteriormente uma pequena cavidade do manto.
Informações sobre crescimento e maturidade sexual são escassas. Sabe-se que espécies de Prochaetoderma youngei atingem o tamanho adulto e torno dos 2 meses e sua maturidade sexual em 1 ano. Estas são informações de animais mantidos em tanques, já que a espécie vive em profundidades em torno de 2000 metros.
Rios(1994) informa a existência de 4 espécies no brasil: Neomenia herwigi, Falcidens sp., Scutopus cf. megaradulatus e Limifossor sp.
A classe está dividida em dois grupos: Solenogastres (Neomeniomorpha) e Caudofoveata (Chaetodermomorpha).
Solenogastres: possuem uma abertura ventral estreita onde está o pé, que não passa de uma protuberância ciliada sem músculos por onde se arrasta. São hermafroditas.
Caudofovetas: diferentemente dos Solenogastres estes não possuem um pé ou abertura ventral. São dióicos (sexos separados). A cabeça é geralmente separada do corpo por uma pequena constrição.
Ecologia
Esses animais podem ser encontrados desde a zona de marés até profundidades em torno de 9000 metros. Parecem ser bastante abundantes, embora sejam pouco estudados por viverem em águas profundas. Algumas espécies ocorrem em todo o mundo e algumas espécies até dominam a macro fauna local. Ocorrem desde o Ártico até a Antártida. Vivem em fundos lodosos, sobre hidróides ou octocorais.
Estrutura
São animais que variam de 1mm até 30 cm. Seus corpos variam do quase esférico, alongado, achatado e estreito.
Alimentação
São animais carnívoros e/ou ornívoros que se alimentam de foraminíferos, detritos e cnidários.
Classe Scaphopoda
SCAPHOPODA (do grego skaphe, pá + pous, podos, pé – em referência ao pé, que o molusco utiliza para escavar e se enterrar no substrato).
É a menor classe dentre os moluscos com algo em torno de 550 espécies em todo o mundo. No Brasil são conhecidas aproximadamente 30 espécies.
Os escafópodes são moluscos marinhos caracterizados por possuirem uma concha tubular e aberta nas duas extremidades. Em muitas espécies, como por exemplo nos Dentaliidae, as conchas são cônicas, muito alongadas e levemente recurvadas, semelhantes a diminutas presas de elefantes: daí a denominação inglesa “tusk-shells”. As partes moles ficam confinadas no interior da concha. O comprimento da concha varia de 3 a cerca de 150 mm. As conchas de muitas espécies são transparentes ou brancas, mas existem espécies com conchas esverdeadas, amareladas, castanhas, róseas ou avermelhadas. Podem ser lisas, reticuladas, ou com estrias ou carenas longitudinais ou transversais.
A posição filogenética do grupo é bastante controversa. Por exemplo, Brusca & Brusca (2003) e Simone (2009) consideram os bivalves mais relacionados filogeneticamente com os escafópodes do que com os demais moluscos. Já Giribet et al. (2006), com base exclusivamente em dados moleculares, considera os escafápodes mais relacionados com os solenogastres (moluscos desprovidos de conchas) e com os cefalópodes.
Os escafópodes são bentônicos e vivem com a extremidade mais larga da concha, onde se situam o pé e a cabeça rudimentar, enterrada no sedimento, enquanto que a porção mais afilada da concha se projeta para fora do substrato. Os escafópodes utilizam o pé, desprovido de olhos, cilíndrico e ponteagudo, para escavar. A cabeça possui projeções em forma de tentáculos retráteis, denominados captáculos. Os captáculos são ciliados e atuam como orgão tácteis e adesivos para apreensão do alimento, o qual pode ser constituído de detritos existentes no sedimento ou de pequenos organismos, como foraminíferos ou pequenos bivalves. Possuem rádula bem desenvolvida, responsável por moer o alimento capturado. Pelo orifício da porção afilada da concha, circulam as correntes de água responsáveis pela respiração e pela eliminação de resíduos do metabolismo.
Os adultos possuem sexos separados. As fêmeas liberam os ovos um por vez. O desenvolvimento é semelhante ao de vários outros moluscos. Do ovo eclode uma larva trocófora, que se transforma em um véliger. Após algum tempo nadando no plâncton, o véliger se dirige ao fundo, onde se transforma no adulto. Os adultos podem ser encontrados em fundos inconsolidados, lodo e areia, desde regiões de águas rasas até 4.755m de profundidade.
Segundo Brusca & Brusca (2003), existem cerca de 900 espécies descritas de escafópodes. De acordo com o “site” Malacolog, 101 espécies de Scaphopoda estão registradas para o Atlântico Oeste. Rios (1994) relacionou 30 espécies para o Brasil. Caetano & Absalão (2005) descreveram uma espécie nova de Polyschides, e Caetano et.al. (2006) descreveram mais duas espécies novas de Cadulus, registraram quatro novas ocorrências de espécies de escafópodes para o litoral brasileiro, e propuseram três novas combinações genéricas para espécies já conhecidas do Brasil. Caetanoe& Scarabino (in Rios, 2009) registram 34 espécies para o litoral brasileiro.
Morfologia da concha
Morfologia do animal
Ecologia
Classe Bivalvia

Bivalves, também chamados de Lamellibranchia ou Pelecypoda, são a segunda maior Classe de moluscos com aproximadamente 15.000 espécies. São um grupo exclusivamente aquático, bilateralmente simétricos, caracterizados por um corpo comprimido lateralmente com uma concha externa composta por duas valvas. Algumas espécies contudo, como as que vivem presas a algum substrato, como as ostras, não são bilateralmente simétricas e umas poucas possuem uma concha interna.
A concha pode ser totalmente ou parcialmente calcificada e consiste de uma valva esquerda e uma direita. As valvas são unidas na parte dorsal, conectadas por uma ligamento elástico parcialmente calcificado e mantida juntas por músculos adutores, um ou dois, que estão presos à superfície interna da concha. As valvas são abertas pelo ligamento e fechadas pela contração dos músculos adutores.
Sua anatomia diverge consideravelmente do plano básico de um molusco, por não possuir cabeça ou rádula.
A maior parte dos bivalves é filtradora, mas também podem ser necrófagos (carniceiros) e até predadores.
Vivem principalmente enterrados em substratos arenosos ou lodosos, podendo movimentar-se com o uso de seu pé, embora não seja tão eficiente como o dos gastrópodes. Outros como as ostras e mexilhões vivem presos a substratos sólidos e outros podem penetrar em madeira ou até em pedras.
Alguns Pectinides são um exemplo de bivalves que podem nadar por curtas distâncias, fechando rapidamente suas valvas, o que causa deslocamento de água que o impulsiona promovendo uma fuga rápida de predadores.


Escultura externa:
As conchas dos bivalves antigos possuíam muito pouca escultura externa. O desenvolvimento de costelas, escamas e espinhos apareceram nas espécies que passaram a habitar águas mais rasas e quentes, que foi interpretado como mecanismo de defesa ou para estabilização da concha. Os tipos de escultura ilustrados, estão na sua forma mais simples, mas uma combinação destes elementos forma várias espécies de nossa fauna.

Tipos de musculatura
O sistema muscular consiste principalmente de músculos adutores, pé, músculos do manto e os sifões. Os músculos adutores conectam as valvas esquerda e direita. Mesmo os moluscos que possuem um único músculo adutor em fase adulta, já possuíram os dois músculos em alguma fase inicial de sua vida. Quando a concha possui os dois músculos ela é chamada dimiana.

Orientação dos umbos:
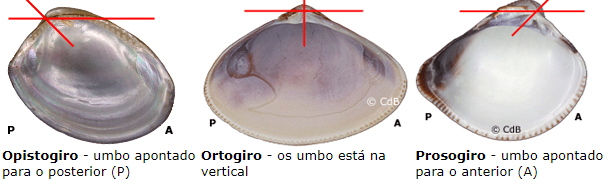
A concha de um bivalve cresce de acordo com uma curva logarítmica, que permite um aumento do volume. O crescimento de uma concha pode ocorrer apenas ao longo da margem ventral, sendo assim, principalmente radial. Diferenças na taxa de crescimento relativo determinam a convexidade da concha. Como em algumas espécies este crescimento não é simétrico, resulta em um menor crescimento em uma das partes, causando um giro na axial do umbo, apontando-o para o posterior ou anterior.
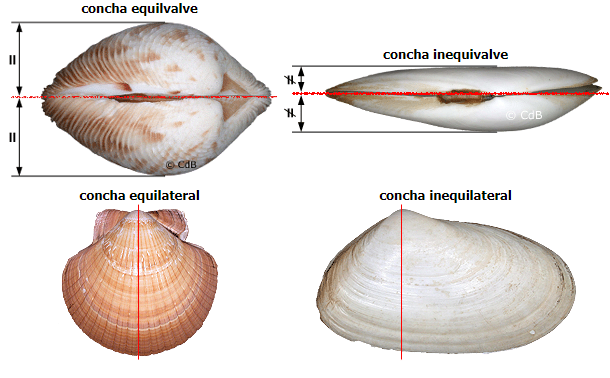
Simetria das valvas:
As conchas que possuem as valvas direita e esquerda de igual tamanho são chamadas equivalves, já conchas de famílias como Mesodesmatides e Pteriides que possuem valvas de tamanhos diferentes são chamados inequivalves. Os Pectinides são tipicamente equilaterais por possuírem o umbo no centro da distância entre o posterior e o anterior. A maioria dos bivalves é inequilateral, onde o umbo está deslocado para o posterior ou anterior, como os Mytilides.